
3.0: Some Applications Leading to Differential Equation
- Page ID
- 15012
This page is a draft and is under active development.

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Much of calculus is devoted to learning mathematical techniques that are applied in later courses in mathematics and the sciences; you wouldn't have time to learn much calculus if you insisted on seeing a specific application of every topic covered in the course. Similarly, much of this section is devoted to methods that can be applied in later courses. Only a relatively small part of the section is devoted to the derivation of specific differential equations from mathematical models or relating the differential equations that we study to specific applications. In this section, we mention a few such applications.
The mathematical model for an applied problem is almost always simpler than the actual situation being studied, since simplifying assumptions are usually required to obtain a mathematical problem that can be solved. For example, in modelling the motion of a falling object, we might neglect air resistance and the gravitational pull of celestial bodies other than Earth, or in modeling population growth we might assume that the population grows continuously rather than in discrete steps.
A good mathematical model has two important properties:
1. It's sufficiently simple so that the mathematical problem can be solved.
2. It represents the actual situation sufficiently well so that the solution to the mathematical problem predicts the outcome of the real problem to within a useful degree of accuracy. If results predicted by the model don't agree with physical observations, the underlying assumptions of the model must be revised until satisfactory agreement is obtained.
We'll now give examples of mathematical models involving differential equations. We'll return to these problems at the appropriate times, as we learn how to solve the various types of differential equations that occur in the models.
All the examples in this section deal with functions of time, which we denote by \(t\). If \(y\) is a function of \(t, y'\) denotes the derivative of \(y\) with respect to \(t\); thus,
\begin{eqnarray*}
y'={dy\over dt}
\end{eqnarray*}
Population Growth and Decay
Although the number of members of a population (people in a given country, bacteria in a laboratory culture, wildflowers in a forest, etc.) at any given time \(t\) is necessarily an integer, models that use differential equations to describe the growth and decay of populations usually rest on the simplifying assumption that the number of members of the population can be regarded as a differentiable function \(P=P(t)\). In most models, it is assumed that the differential equation takes the form
\[P' = a(P)P \label{3.1.1}\]
where \(a\) is a continuous function of \(P\) that represents the rate of change of population per unit time per individual. In the \( \textcolor{blue}{\mbox{Malthusian model}} \) (https://en.Wikipedia.org/wiki/Thomas_Robert_Malthus), it is assumed that \(a(P)\) is a constant, so equation \ref{3.1.1} becomes
\[P' = aP. \label{3.1.2}\]
This model assumes that the numbers of births and deaths per unit time are both proportional to the population. The constants of proportionality are the \( \textcolor{blue}{\mbox{birth rate}} \) (births per unit time per individual) and the \( \textcolor{blue}{\mbox{death rate}} \) (deaths per unit time per individual); \(a\) is the birth rate minus the death rate. You learned in calculus that if \(c\) is any constant then
\[P = ce^{at} \label{3.1.3}\]
satisfies Equation \ref{3.1.2}, so Equation \ref{3.1.2} has infinitely many solutions. To select the solution of the specific problem that we are considering, we must know the population \(P_0\) at an initial time, say \(t = 0\). Setting \(t = 0\) in Equation \ref{3.1.3} yields \(c = P(0) = P_0\), so the applicable solution is
\[P(t) = P_0e^{at}.\nonumber \]
This implies that
\[\lim_{t\to\infty}P(t)=\left\{\begin{array}{cl}\infty&\mbox{ if }a>0,\\ 0&\mbox{ if }a<0; \end{array}\right.\nonumber\]
that is, the population approaches infinity if the birth rate exceeds the death rate, or zero if the death rate exceeds the birth rate.
To see the limitations of the Malthusian model, suppose we're modeling the population of a country, starting from a time \(t=0\) when the birth rate exceeds the death rate (so \(a>0\)), and the country's resources in terms of space, food supply, and other necessities of life can support the existing population. Then the prediction \(P=P_0e^{at}\) may be reasonably accurate as long as it remains within limits that the country's resources can support. However, the model must inevitably lose validity when the prediction exceeds these limits. (If nothing else, eventually there won't be enough space for the predicted population!)
This flaw in the Malthusian model suggests the need for a model that accounts for limitations of space and resources that tend to oppose the rate of population growth as the population increases. Perhaps the most famous model of this kind is the \( \textcolor{blue}{\mbox{Verhulst model}} \) (http://www-history.mcs.st-and.ac.uk/.../Verhulst.html) where equation \ref{3.1.2} is replaced by
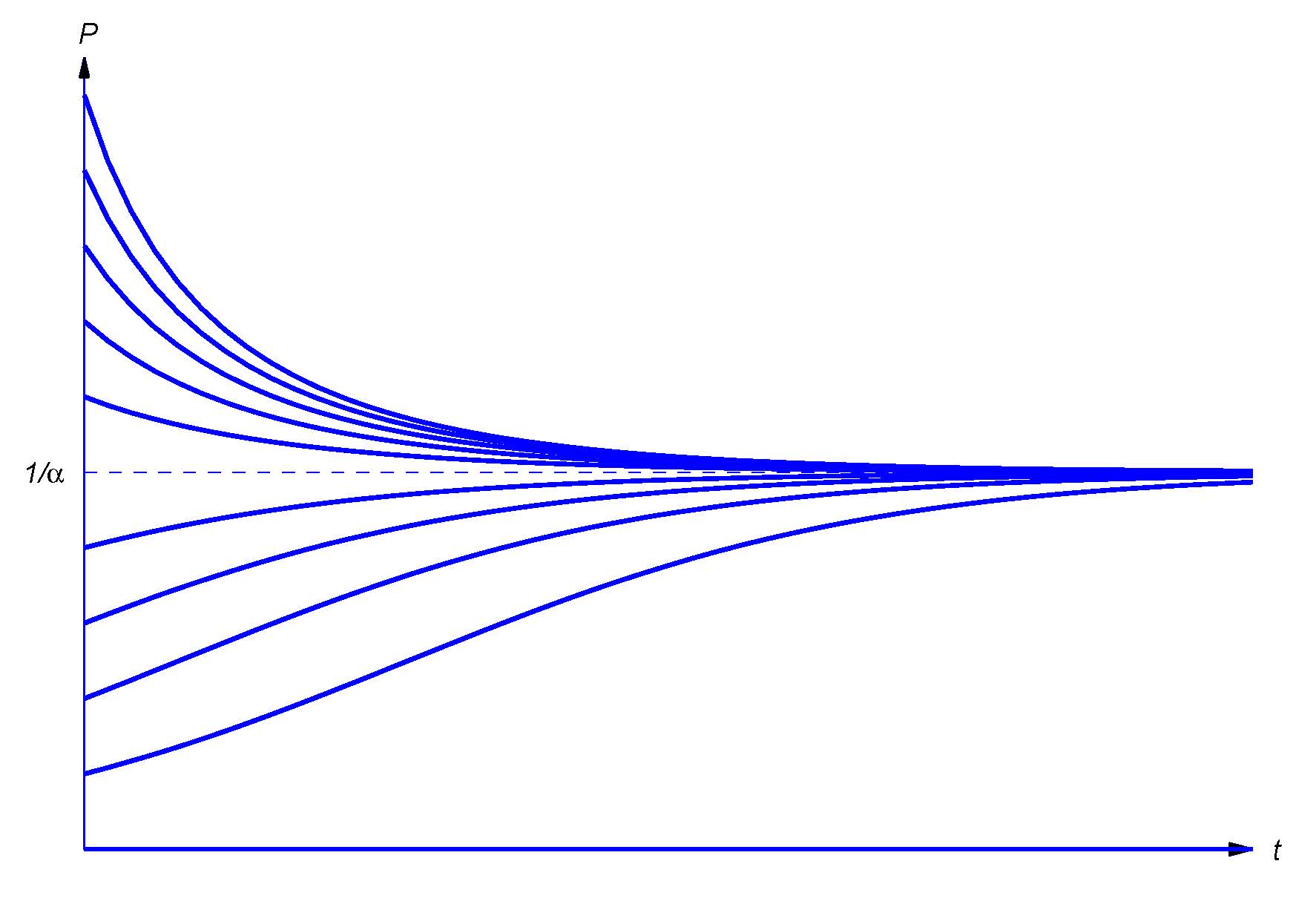
\[P'=aP(1-\alpha P), \label{3.1.4} \]
where \(\alpha\) is a positive constant. As long as \(P\) is small compared to \(1/\alpha\), the ratio \(P'/P\) is approximately equal to \(a\). Therefore the growth is approximately exponential; however, as \(P\) increases, the ratio \(P'/P\) decreases as opposing factors become significant.
Equation \ref{3.1.4} is the \( \textcolor{blue}{\mbox{logistic equation}} \). You will learn how to solve it in Section 3.2. (See Exercise 3.2.28.) The solution is
\[P={P_0\over\alpha P_0+(1-\alpha P_0)e^{-at}} \]
where \(P_0=P(0)>0\). Therefore \(\lim_{t\to\infty}P(t)=1/\alpha\), independent of \(P_0\).
Figure \(3.1.1\) shows typical graphs of \(P\) versus \(t\) for various values of \(P_0\).
Newton's Law of Cooling
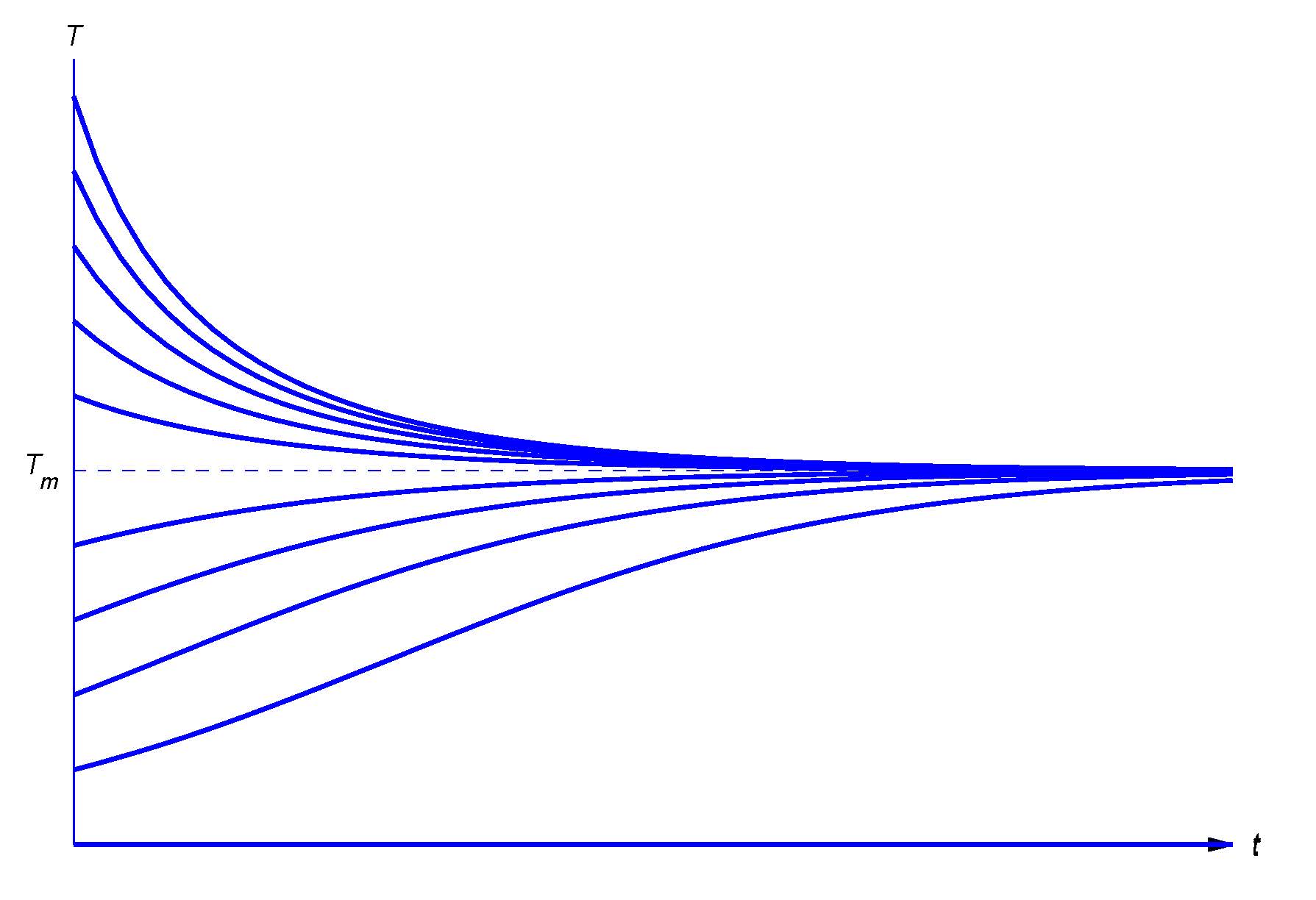
According to \( \textcolor{blue}{\mbox{Newton's law of cooling}} \) (http://www-history.mcs.st-and.ac.uk/...es/Newton.html), the temperature of a body changes at a rate proportional to the difference between the temperature of the body and the temperature of the surrounding medium. Thus, if \(T_m\) is the temperature of the medium and \(T=T(t)\) is the temperature of the body at time \(t\), then
\[T_0 = −k(T −T_m) \label{3.1.5}\]
where \(k\) is a positive constant and the minus sign indicates; that the temperature of the body increases with time if it's less than the temperature of the medium, or decreases if it's greater. We'll see in Section~3.2 that if \(T_m\) is constant then the solution of equation \ref{3.1.5} is
\[T = T_m + (T_0 −T_m)e^{−kt} \label{3.1.6}\]
where \(T_0\) is the temperature of the body when \(t=0\). Therefore \(\lim_{t\to\infty}T(t)=T_m\), independent of \(T_0\). (Common sense suggests this. Why?)
Figure \(3.1.2\) shows typical graphs of \(T\) versus \(t\) for various values of \(T_0\).
Assuming that the medium remains at constant temperature seems reasonable if we're considering a cup of coffee cooling in a room, but not if we're cooling a huge cauldron of molten metal in the same room. The difference between the two situations is that the heat lost by the coffee isn't likely to raise the temperature of the room appreciably, but the heat lost by the cooling metal is. In this second situation, we must use a model that accounts for the heat exchanged between the object and the medium. Let \(T=T(t)\) and \(T_m=T_m(t)\) be the temperatures of the object and the medium respectively, and let \(T_0\) and \(T_{m0}\) be their initial values. Again, we assume that \(T\) and \(T_m\) are related by equation \( (5) \). We also assume that the change in heat of the object as its temperature changes from \(T_0\) to \(T\) is \(a(T-T_0)\) and the change in heat of the medium as its temperature changes from \(T_{m0}\) to \(T_m\) is \(a_m(T_m-T_{m0})\), where \(a\) and \(a_m\) are positive constants depending upon the masses and thermal properties of the object and medium respectively. If we assume that the total heat of the in the object and the medium remains constant (that is, energy is conserved), then
\[a(T −T_0) + a_m(T_m −T_{m0}) = 0. \nonumber\]
Solving this for \(T_m\) and substituting the result into Equation \ref{3.1.6} yields the differential equation
\[T ^ { \prime } = - k \left( 1 + \frac { a } { a _ { m } } \right) T + k \left( T _ { m 0 } + \frac { a } { a _ { m } } T _ { 0 } \right) \nonumber\]
for the temperature of the object. After learning to solve linear first order equations, you’ll be able to show (Exercise 4.2.17) that
\[T = \frac { a T _ { 0 } + a _ { m } T _ { m 0 } } { a + a _ { m } } + \frac { a _ { m } \left( T _ { 0 } - T _ { m 0 } \right) } { a + a _ { m } } e ^ { - k \left( 1 + a / a _ { m } \right) t }\nonumber \]
Glucose Absorption by the Body
Glucose is absorbed by the body at a rate proportional to the amount of glucose present in the bloodstream. Let \(\lambda\) denote the (positive) constant of proportionality. Suppose there are \(G_0\) units of glucose in the bloodstream when \(t=0\), and let \(G=G(t)\) be the number of units in the bloodstream at time \(t>0\). Then, since the glucose being absorbed by the body is leaving the bloodstream, \(G\) satisfies the equation
\[G_0 = −λG. \label{3.1.7}\]
From calculus you know that if \(c\) is any constant then
\[G = ce^{−λt} \label{3.1.8}\]
satisfies Equation 3.0.8, so Equation \ref{3.1.8} has infinitely many solutions. Setting \(t = 0\) in Equation \ref{3.1.8} and requiring that \(G(0) = G_0\) yields \(c = G_0\), so
\[G(t) = G_0e^{−λt}.\nonumber \]
Now let's complicate matters by injecting glucose intravenously at a constant rate of \(r\) units of glucose per unit of time. Then the rate of change of the amount of glucose in the bloodstream per unit time is
\[G' = −λG + r \label{3.1.9}\]
where the first term on the right is due to the absorption of the glucose by the body and the second term is due to the injection. After you've studied Section~3.1, Exercise~3.2.8, you’ll be able to show that the solution of Equation \ref{3.1.9} that satisfies \(G(0) = G_0\) is
\[G = \frac { r } { \lambda } + \left( G _ { 0 } - \frac { r } { \lambda } \right) e ^ { - \lambda t }\nonumber \]
Graphs of this function are similar to those in Figure \(1.1.2\). (Why?)
Spread of Epidemics
One model for the spread of epidemics assumes that the number of people infected changes at a rate proportional to the product of the number of people already infected and the number of people who are susceptible, but not yet infected. Therefore, if \(S\) denotes the total population of susceptible people and \(I=I(t)\) denotes the number of infected people at time \(t\), then \(S-I\) is the number of people who are susceptible, but not yet infected. Thus,
\begin{eqnarray*}
I'=rI(S-I),
\end{eqnarray*}
where \(r\) is a positive constant. Assuming that \(I(0)=I_0\), the solution of this equation is
\[I =\dfrac{SI_0}{I_0 + (S −I_0)e^{−rSt}}\nonumber \]
(Exercise~3.4.29).
Graphs of this function are similar to those in Figure \(1.1.1\). (Why?)
Since \(\lim_{t\to\infty}I(t)=S\), this model predicts that all the susceptible people eventually become infected.
Newton's Second Law of Motion
According to \( \textcolor{blue}{\mbox{Newton's second law of motion}} \) (http://www-history.mcs.st-and.ac.uk/...es/Newton.html), the instantaneous acceleration \(a\) of an object with constant mass \(m\) is related to the force \(F\) acting on the object by the equation \(F=ma\). For simplicity, let's assume that \(m=1\) and the motion of the object is along a vertical line. Let \(y\) be the displacement of the object from some reference point on Earth's surface, measured positive upward. In many applications, there are three kinds of forces that may act on the object:
(a) A force such as gravity that depends only on the position \(y\), which we write as \(-p(y)\), where \(p(y)>0\) if \(y\ge0\).
(b) A force such as atmospheric resistance that depends on the position and velocity of the object, which we write as \(-q(y,y')y'\), where \(q\) is a nonnegative function and we've put \(y'\) "outside'' to indicate that the resistive force is always in the direction opposite to the velocity.
(c) A force \(f=f(t)\), exerted from an external source (such as a towline from a helicopter) that depends only on \(t\).
In this case, Newton’s second law implies that
\[y'' = −q(y,y')y' −p(y) + f(t), \nonumber\]
which is usually rewritten as
\[y'' + q(y,y')y' + p(y) = f(t). \nonumber\]
Since the second (and no higher) order derivative of \(y\) occurs in this equation, we say that it is a second order differential equation.
Interacting Species: Competition
Let \(P=P(t)\) and \(Q=Q(t)\) be the populations of two species at time \(t\), and assume that each population would grow exponentially if the other didn't exist; that is, in the absence of competition we would have
\[P'=aP \quad \text{and} \quad Q'=bQ, \label{3.1.10}\]
where \(a\) and \(b\) are positive constants. One way to model the effect of competition is to assume that the growth rate per individual of each population is reduced by an amount proportional to the other population, so Equation \ref{3.1.10} is replaced by
\[\begin{align*} P' &= aP-\alpha Q\\[4pt] Q' &= -\beta P+bQ,\end{align*}\]
where \(\alpha\) and \(\beta\) are positive constants. (Since negative population doesn't make sense, this system works only while \(P\) and \(Q\) are both positive.) Now suppose \(P(0)=P_0>0\) and \(Q(0)=Q_0>0\). It can be shown (Exercise~3.4.28) that there's a positive constant \(\rho\) such that if \((P_0,Q_0)\) is above the line \(L\) through the origin with slope \(\rho\), then the species with population \(P\) becomes extinct in finite time, but if \((P_0,Q_0)\) is below \(L\), the species with population \(Q\) becomes extinct in finite time. Figure \(3.1.3\) illustrates this. The curves shown there are given parametrically by \(P=P(t), Q=Q(t),\ t>0\). The arrows indicate direction along the curves with increasing \(t\).

Contributors
Trench, William F., "Elementary Differential Equations" (2013). Faculty Authored and Edited Books & CDs. 8.
https://digitalcommons.trinity.edu/mono/8